发布时间:2021-09-03
——《理想国》
每个瞬间,我们都被无数的外界刺激所包裹。窗外的鸟鸣、屏幕上的文字、远处飘过来的咖啡香、走廊上响起的脚步声……即使是一小捧清水中,也包含着巨大的信息量。
而同时,我们很少能够获取关于某个事物的全部信息。我们看到迎面飞过来的羽毛球,却无法“看到”它完整的运动轨迹;我们听到远处的雷声,却不知道大雨何时会从头顶落下;我们看到朋友脸上的表情,却不能直接看到他此时此刻的心情。
我们所能感知到的世界,一如被困在洞穴中的人所能看到的石壁上的影子。这些由我们所接收到的感觉刺激所组成的瞬间,只是世界的切片。但我们却能够在茫茫人海中,一眼看到自己熟悉的面孔,也能够借由一片落叶而知晓秋天的到来。这是由于我们对环境的感知,并不只是被动地接受刺激,而是一个主动对这些刺激进行加工和诠释的过程。就像为了尽可能准确描述影子所对应的实物,我们一方面需要从它的众多特征中选取真正重要的那些,例如它的长宽高是重要的,而石壁本身的质地则相对不那么重要;另一方面,还需要对一些不能直接观察到的变量进行推测,例如实物距离石壁的距离,火光所在的方向。这两个过程,都受到过往的经验所影响。在经验的引导下,我们能建立起一个自己的坐标系,用以描述眼前的一切。
而我们之所以能够建立这样一个坐标系,是因为大脑对感觉刺激的表征本身就受到各种因素的调控。即使对于同一感觉刺激而言,在学习前后,大脑对于该刺激的反应也会有所不同[1]。而同时,对感觉刺激的编码在不同状态下也会有所不同。例如,相对于静止状态,在运动状态下听觉皮层的活动会有所减弱[2]。这些差异的存在是由于感觉皮层的神经元接收到了来自下游脑区的反馈投射。这些编码了复杂信息的反馈投射活动,调节了感觉皮层在不同条件下的反应强度[3],就像一面哈哈镜,随时随地调节着对于外界影像的反映。
感觉皮层中不仅存在对感觉刺激本身的编码,还编码了很多重要的隐含变量。在一个分类任务中,如果被试需要将所接受到的听觉刺激分成“高频”和“低频”两组。那么,在这个过程中会有相当数量的感觉皮层的神经元表现出对于类别的编码[4]。这也许就是为何即使光谱是连续的,我们依旧用“赤橙黄绿蓝靛紫”这样一些离散的颜色来描述彩虹。
这种对类别的编码,跳脱出了对于刺激特性一对一的固化的表征形式,使得我们可以更加有效地从复杂的、高维的环境信息中提取出与真正与当前所面临的行为选择相关的特征。

图1. 大脑能对接收到的感觉刺激进行分类(改编自Kourtzi and Connor, 2011[5])
广泛存在的演绎法
“即使没有亲眼目睹、亲耳听说大西洋或者尼亚加拉大瀑布,逻辑学家也可以由一滴水推断出二者的存在。生活实则是根巨大的链条,观其一环便可知整体。”
——《福尔摩斯探案集》
我们常说“举一反三”、“窥一斑而知全豹”。这样一种泛化和推测的能力,是大脑经过亿万年演化后才具备的神奇能力。
早在1949年,Harlow就在人类和猕猴上的行为学实验发现,被试能够通过学习发现潜在的任务规则,作出泛化,从而更加高效地完成新任务。在这一实验中,每次会有两个视觉刺激被呈现给被试,而其中,只有一个刺激与奖励相关联,被试需要选择该刺激从而获取奖励。每六次测试之后,新的一组视觉刺激会替换掉原有的一组,其中依然只有一个刺激与奖赏相关。实验发现,随着经验的增加,在面对新的感官刺激时,被试的学习速度会逐渐加快[6]。这表明,在整个实验的过程中,被试不仅是在单纯地学习哪一个特定刺激与奖励相连,而是学习到了一些与任务结构相关的信息,即“两个图片中总有一个图片对应奖励”这一规则,这使得其后期的学习变得更为高效。
我们所身处的这个世界虽然复杂多变,但在每个抽象层次上都充满着重复性和自我相似性。正因如此,不同的外界感觉刺激也许会预示着相同的信息,从而介导了相似的行为策略。比如我们上文所描述的关于分类的例子,虽然具体的声音刺激频率不同,但它们都可以按照一定的规则被分到“高频”的类别中。这些结构特征的存在也意味着,即使在一个崭新的情景下,原有的经验依旧可以对当下的决策给予指导。
生物体如果建立起一个有关环境因果关系的内在模型,便能够在接收到较少反馈的情况下,较为快速地理解环境的变化从而做出恰当的行为选择。那么,这样一种内在模型在我们的大脑中是否存在,又会以怎样的形式被编码呢?
大脑中的图书馆
“图书馆是无限的,周而复始的。”
——《巴别塔图书馆》
博尔赫斯笔下的宇宙,是一个由不断重复的六角形回廊组成的图书馆。这样一种自我重复的结构与我们对于真实世界的表征相似。
如果在我们的大脑中真的存在这样一个图书馆,那么构成这个图书馆的神经元或者神经元集群的活动应该满足两个条件:表征了环境中与行为选择相关的重要变量;这种表征能够在具有相似结构的环境中进行泛化。
目前的研究中,内嗅皮层-海马系统、眶额叶,以及内侧前额叶被认为具有这样的信息编码特性。这些脑区的神经元活动往往不依赖具体的感觉刺激,而是在不同实验中都对某一任务相关特征进行稳定编码[7]。同时,这些脑区中信息编码的侧重点又有所不同。内嗅皮层和内侧前额叶通常被认为编码了更加抽象的结构信息;海马中对结构信息的编码会随着具体任务的改变而有所不同;与这些区域对于结构信息的编码不同,眶额叶的神经元则似乎主要反映了那些无法直接通过观察得到的隐含变量,例如已经做出的行为选择、得到过的外界反馈、对于奖赏的期待等。
每个脑区中所编码的信息就像图书馆中的一层,它们分别代表了信息检索中某一个重要的环节,通过相互连接而表征了完整的信息链。这种连接由脑区间神经元的相互投射完成。
一些理论研究认为,海马同时接收来自内嗅皮层的抽象的结构信息和来自其他脑区的感觉信息,并对这二者进行联合编码[8]。这就像在进化树的每个节点上,加上具体的物种。
而解剖学的证据表明,眶额叶和各种感觉皮层以及包括腹侧被盖区、杏仁核在内的在学习过程中扮演着重要角色的脑区之间都具有紧密的投射关系[9]。通过抑制或是损毁的手段改变眶额叶皮层神经元的活动,也会使下游脑区的神经元活动发生改变,从而影响学习、抉择灵活性等行为过程。
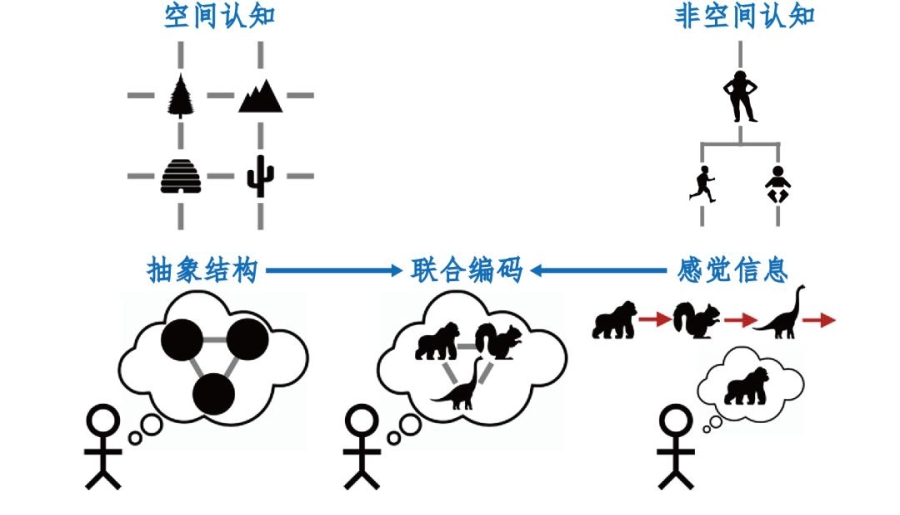
图2. 大脑中的认知地图(改编自Whittington et al., 2020[8])
这些对内在模型进行编码的脑区,与编码了感觉刺激、奖赏反馈的区域相互作用,使得即使在信息有限、充满噪声的世界中,生物依旧能够对自身所处的环境做出快速的判断、从而做出恰当的行为选择。
然而,大脑对内在模型的表征和运用依然存在很多尚未解决的问题。对于这一过程的研究本身,也是一个试图通过有限的信息、充满噪声的实验结果去描述大脑的过程。与试图通过影子去判断火光背后的实体异曲同工。希望有一天,我们也能像我们的大脑一样,在日复一日的经验积累下、在每一次或对或错的尝试中逐渐寻找到事件与事件之间的因果关系,建立起一个恰当的坐标系,用以描述我们自己。
参考文献:
[1] D. B. Polley, “Perceptual Learning Directs Auditory Cortical Map Reorganization through Top-Down Influences,” J. Neurosci., vol. 26, no. 18, pp. 4970–4982, 2006.
[2] D. M. Schneider, A. Nelson, and R. Mooney, “A synaptic and circuit basis for corollary discharge in the auditory cortex,” Nature, vol. 513, no. 7517, pp. 189–194, 2014.
[3] Y. Liu, Y. Xin, and N. Xu, “A cortical circuit mechanism for structural knowledge-based flexible sensorimotor decision-making,” Neuron, pp. 1–16, 2021.
[4] Y. Xin, L. Zhong, Y. Zhang, T. Zhou, J. Pan, and N. long Xu, “Sensory-to-Category Transformation via Dynamic Reorganization of Ensemble Structures in Mouse Auditory Cortex,” Neuron, vol. 103, no. 5, pp. 909-921.e6, 2019.
[5] Z. Kourtzi and C. E. Connor, “Neural Representations for Object Perception: Structure, Category, and Adaptive Coding,” Annu. Rev. Neurosci., vol. 34, no. 1, pp. 45–67, Jul. 2011.
[6] H. F. Harlow, “The formation of learning sets,” Psychol. Rev., vol. 56, no. 1, pp. 51–65, Jan. 1949.
[7] A. M. Wikenheiser and G. Schoenbaum, “Over the river, through the woods: cognitive maps in the hippocampus and orbitofrontal cortex,” Nat. Rev. Neurosci., vol. 17, no. 8, pp. 513–523, Aug. 2016.
[8] J. C. R. Whittington et al., “The Tolman-Eichenbaum Machine: Unifying Space and Relational Memory through Generalization in the Hippocampal Formation,” Cell, vol. 183, no. 5, pp. 1249-1263.e23, 2020.
[9] B. Zingg et al., “Neural Networks of the Mouse Neocortex,” Cell, vol. 156, no. 5, pp. 1096–1111, Feb. 2014.
 附件下载:
附件下载: 