发布时间:2020-11-09
高三的表妹最近记性不太好,今天背的课文明天就忘,为此非常头疼,常常向我吐槽。一天她突发奇想地问我能不能把这种情况反转一下,也就是说今天背的东西今天先忘记掉,等到考试那天再回想起来。
和大多数读者一样,笔者对这个想法莞尔而笑,但仔细一想,却发觉这并不违反基本的科学原理。
表妹的问题在于短期记忆不错,能记住今天背的课文;但长期记忆欠佳,过了一段时间就忘记了。而她希望的状态则与现状相反,盼望能将自己的短期记忆和长期记忆互换,即虽然今天背的课文今天记不住,但是随着时间的推移,长期记忆将慢慢巩固,到考试那天就能下笔如有神了。
关键问题就在于,长期记忆是否需要短期记忆作为基础?或者说短期记忆是否是通向长期记忆的一个中间步骤?抑或长期记忆和短期记忆是两个相对独立的过程,可以平行运作?
这个问题的答案其实并不是显而易见的,不同的理论和学派至少争论了有半个多世纪。
Part. 1
大鼠实验:揭开记忆平行宇宙的面纱
根据记忆持续的时间尺度可以粗略地将其分为仅仅持续数秒到若干小时的短期记忆(short-term memory)以及更长时间尺度的长期记忆。也有人将长期记忆继续细分,其中一部分持续长达若干月,甚至铭记终身的被称作持久记忆(long-lasting memory),而较短的部分则是研究较多的所谓长期记忆(long-term memory)。
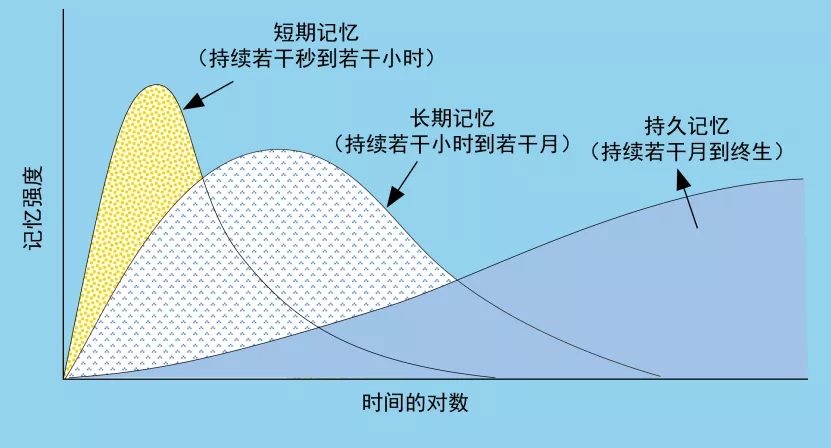
图表 1 记忆巩固的阶段(图片改编自参考文献(1))
于上世纪中叶提出的双痕迹假说(dual-trace hypothesis)认为长期记忆起源于短期记忆(1)。后者的潜在机制是神经元间循环往复的电活动交流,而这些活动固化下来就变成了长期记忆。很多实验的结果与此假说一致,如一些药物可以影响长期记忆,但不影响短期记忆。
不过后来也渐渐出现了一些与之相悖的实验结果。例如有人用伸脚抑制性回避实验来检测大鼠的长期和短期记忆。

图表 2 伸脚抑制性回避实验。训练时大鼠放在高台上,下地就会受到电击。测试时它如果记得被电击的不愉快经历则会不情愿下地或者下地速度减慢。图片来源:作者改编
具体来说,就是将大鼠放在一个高而狭小的台子上,台子下面是铺了金属网格的地面。由于台子过小,大鼠站立不稳就会慢慢伸脚下探到地面。当它四脚落在金属网格上时,会受到网格的电击,这对它是一个负面的刺激。经过不同的时间之后再次将大鼠放在小台子上,这时可以通过记录它四脚落地的时间来量化大鼠的短期记忆(1.5小时之后测量)和长期记忆(24小时之后测量)。
因为如果大鼠记得电击的经历,会更加不愿意下降到地面,就会下降得更加缓慢。下降得越快说明其遗忘得越多,反之下降得越慢则说明其记住得越多。
研究发现,如果在大鼠落地接受电击以后,马上在其海马体或内嗅皮层(大脑中与记忆相关的部位)注射多种药物(如蝇蕈醇)会产生奇妙的效果,即不影响其短期记忆,而却影响其长期记忆。这些结果提示长期记忆和短期记忆也许不是串联的关系,可能是相对独立的并行关系(2)。
Part. 2
海兔实验:不同记忆的形成机理是怎样的?
为了深入探究长期记忆与短期记忆的机理,生物学家坎德尔选中了海兔作为研究对象,因为相比于人脑有上千亿个神经元,海兔仅有2万,而且和学习直接相关的中枢神经细胞不超过100个。
除此以外,海兔还拥有其它诸多优势。比如它的细胞很大(直径可达1毫米),可以肉眼观察,也可以直接注入多种化合物用于标记等。这使得海兔成为了研究学习记忆的一个低门槛的好工具(3)。
海兔的缩鳃反应的敏化作用可以作为一个研究学习记忆的好范式,它能模拟短期记忆(单个电击所致的敏化作用持续时间较短)和长期记忆(多个电击所致的敏化作用持续时间长)。
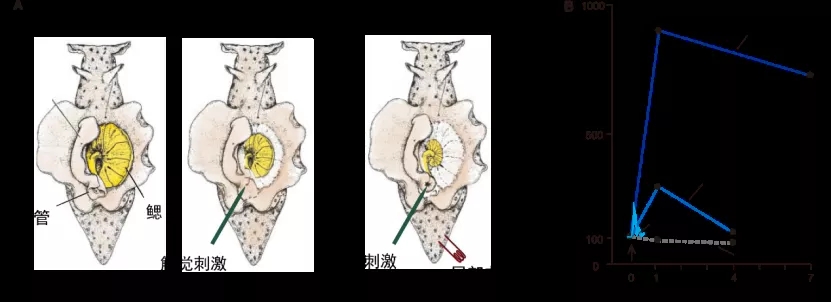
图表 3 一个简单的学习行为。A是海兔背部俯视图,轻触虹吸管会诱导其缩鳃。如果触摸其虹吸管的同时在其尾部施加一个电击,则缩鳃反应会增强,称作敏化作用。B如果反复训练多天,则这种敏化作用会变得更强,并能持续多日。图片来源:改编自参考文献(3)
研究首次发现,长期记忆的形成需要蛋白质合成,而短期记忆不需要蛋白质合成。
在海兔的缩鳃敏化过程中,感觉神经元和其下游运动神经元之间的突触连接发生了强化,使得同样的感觉刺激相同程度地激活了感觉神经元。但是下游的运动神经元可以通过更强的突触接收到更强的上游的输入,从而做出更强烈的行为反应。
因此可以认为,记忆就存储在感觉神经元和运动神经元之间的突触里,只是长期记忆和短期记忆实现的方式有差别。相同之处是他们都需要5-羟色胺这种信号分子,并由此在形成突触的细胞里引发一系列生化反应。
不同点是这些反应有个本质区别:短期记忆只是诱发了钙离子大量涌入感觉神经元时,神经元会倾向于释放更多的递质,从而使突触后运动神经元产生更大的反应。这种过程是暂时的,很快感觉神经元释放递质会恢复正常水平,敏化作用减弱。
反之,如果这些生化反应诱导感觉神经元合成蛋白质,最终以此为原料生成了新的突触,那么从感觉神经元到运动神经元的突触强度就会得到长期增强,直到突触又消亡,这也就是所谓的长期记忆了。
5-羟色胺在此过程中具有双重作用。由于它是在突触附近局部释放,一方面它可以诱导突触传递效率发生变化,产生短期记忆;另一方面当细胞启动新蛋白质合成过程时,5-羟色胺的信号能够利用新合成的蛋白质,标记到底是哪些突触需要增强,从而有针对性地增强这些和新记忆形成相关的突触,或者在相关细胞之间特异性地产生和新记忆相关的新突触。
因此在这个案例中,5-羟色胺才是海兔长期记忆和短期记忆的交汇点,它指示了需要被强化的突触是哪一些,而这是因记忆内容而异的(不同的记忆需要被存放在不同的突触里)。
故大致可以理解为5-羟色胺提供了记忆的内容,其余很多过程都是长期记忆和短期记忆各做各的,平行开展。
Part. 3
人类的记忆更复杂,可以用这种原理解释吗?
当然也有人可能会觉得这个过程太过简单,和人类复杂的记忆现象不可同日而语。

图表 4 陈述性记忆与非陈述性记忆。周末笔者去江边钓鱼,江上百舸争流,半天下来收获颇丰,这个记忆就属于陈述性记忆(如左图)。右图所示钓鱼技能则是非陈述性记忆,因为它不好描述,即使说不出为什么,但是高手一钓一个准。图片来源:作者改编
科学上有两种记忆,一种是陈述性记忆,一种是非陈述性记忆(比如感知和运动技能的训练,不必有意识地回忆过往的事件)。上述海兔的案例属于简单的非陈述性记忆,而更为高级的陈述性记忆一般被认为和海马体中的突触可塑性相关,包括长时程增强和长时程抑制。
这些可塑性分为不需要蛋白质合成的早期和需要蛋白质合成的晚期等若干阶段,其时间尺度分别和短期记忆和长期记忆不谋而合,让人禁不住联想这些突触间的变化和记忆之间的关系。一系列的实验证据支持了这样的猜想。
虽然陈述性记忆和非陈述性记忆从行为表现到内在机制都具有巨大的差异,但是它们都可以分为长期和短期记忆,并且规律惊人地一致,比如:
解释海兔学习记忆的长时程易化(LTF)分为短期过程和长期过程,而解释陈述性记忆的长时程增强(LTP)也可以分为早期和晚期,它们都被分别认为是短期和长期记忆的基础;
短期记忆不需要蛋白质合成(相反,它是通过让上游神经元释放更多递质,以增强连接来实现突触强度改变的),而长期记忆需要蛋白质合成(并且形成新的突触)。
Part. 4
生物为什么要形成不同的记忆?
不过,为什么生物需要同时进行两个不同的过程,分别来实现长期记忆和短期记忆呢?
也许只有当刺激到达一定的强度才能激活一系列信号转导机制,并发动蛋白质合成,产生长期记忆。这时的刺激都是对生物的生存比较重要的。
相反,当刺激不是那么强烈的时候,也许它对生物的生存不那么重要,没必要花费资源去长期记住这些刺激,那么蛋白质合成就未被启动,也就没有形成长期记忆,只形成短期记忆了。
回到文章开头的问题,鉴于长期记忆和短期记忆的机制存在相互独立的部分,如果恰好抑制掉了短期记忆那独有的一部分,则理论上是可能表现出今天背了课文记不住,考试那天往事栩栩如生的状态,只是技术上要实现这种特异性的抑制恐怕还不是一朝一夕的事。
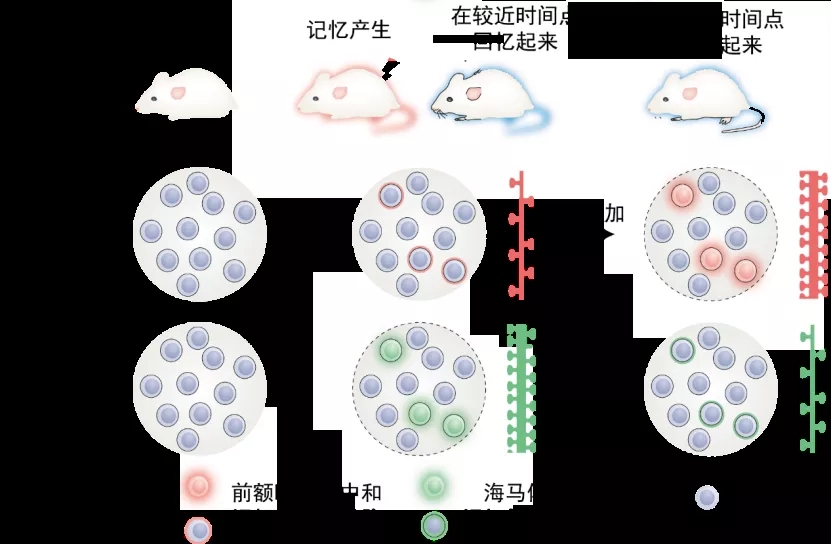
图表 5 长期记忆中前额叶皮层和海马体进行着相反的过程。前额叶皮层中和记忆相关的细胞从不活跃变成活跃状态,突触增加,而海马体中和记忆相关的细胞则从活跃状态变成不活跃状态,并且突触减少。图片来源:改编自参考文献(4)
最后笔者打算讲几句题外话:长期记忆可以分为更细致的阶段,也许分别对应于类似开头提到的长期记忆和持久记忆。在这个转化过程中,人们认为记忆从海马转移到内侧前额叶皮层进行更持久的记忆,一方面海马体内的突触强度在下降,另一方面前额叶皮层的突触强度在上升(4)。看起来好像长期记忆里面也有多个平行宇宙在各司其职。
参考文献:
1. Lechner HA, Squire LR, Byrne JH. 100 Years of Consolidation— Remembering Müller and Pilzecker. Learn Mem. 1999 Mar 1;6(2):77–87.
2. Izquierdo I, Medina JH, Vianna MRM, Izquierdo LA, Barros DM. Separate mechanisms for short- and long-term memory. Behavioural Brain Research. 1999 Aug 1;103(1):1–11.
3. Kandel ER. The Molecular Biology of Memory Storage: A Dialogue Between Genes and Synapses. Science. 2001 Nov 2;294(5544):1030–8.
4. Tonegawa S, Morrissey MD, Kitamura T. The role of engram cells in the systems consolidation of memory. Nat Rev Neurosci. 2018 Aug;19(8):485–98.
 附件下载:
附件下载: 