发布时间:2019-07-03
在普罗大众的印象中,自闭症像是一种性格标签,可以不吝惜地贴在了那些不善社交、少与别人交流、性情内向的人的身上。然而对于医学和神经科学相关的专业人士而言,自闭症却并不是一张“意如其名”的标签,能够轻易贴出来。
自闭症的首次临床定义是在20世纪30-40年代由奥地利医生H.Asperger 与美国医生Kanner共同提出的,经过几十年的发展,医学领域认为自闭症(autism)是一类广泛性的神经系统发育性疾病,包括:瑞特综合症(Rett syndrome)、阿斯伯格综合症(Asperger syndrome)、童年瓦解性障碍等病症。由于是发育性疾病,患者从婴幼年便开始发病,主要的病因是会影响到大脑发育过程的一些基因发生了突变。患者的临床表现主要在:1如对父母不产生依恋,不能与同龄儿童建立伙伴关系等此类社会交往行为的障碍;2如无法用动作表达自己意愿,语言运用和理解能力缺失等此类交流性障碍;3如无法对玩具感兴趣,重复蹦跳,重复机械性动作等此类兴趣缺失及重复刻板性行为,另有少量病患出现癫痫、智力低下、个体发育迟缓等较严重病症。
近几十年来,随着生命科学的日新月异,对于自闭症的研究和理解也越来越深入,从早期的临床病理层面,逐步深入到细胞以及分子的病理层面,使得人们对于自闭症的发生、病程进展以及治疗有了更全面和精细的了解。目前的研究结果显示:自闭症并不是神经系统的严重的、明显的损伤,而是与神经系统的发育过程以及功能具有相关性,其中最重要的则是与神经突触功能与调节息息相关。对于神经系统而言,神经元是此系统的核心,也是最基本的功能单位。神经元彼此之间相互连接才能顺畅地传递神经信号,完成最基本的神经功能。而神经元彼此之间的连接枢纽则构成了大脑行使高级神经活动的基础。神经元间相互连接的界面被称为神经突触,由此可以理解突触对于神经系统的重要性。有趣的是,神经元间的连接并不是静态的,而是一直处于动态的变化当中,旧的不需要的突触消失,新的突触又形成,在这个此消彼长的过程中,消失与形成却不是随机而是处于精密的调控机制操纵之下的。那些与突触这个微小却又重要的结构以及调控这个结构消亡产生相关的生命源代码——基因,毫无疑问,也与自闭症有着密切而重要的联系。
在这些相关的基因中,一个名叫甲基化CpG岛结合蛋白2(MeCP2)的基因,以其有趣的特征吸引了研究者们的注意。MeCP2基因编码一种甲基化DNA结合蛋白,它本身虽然不直接参与到神经突触的结构和功能中,但却可以通过结合DNA的甲基化CpG岛或是“招募”转录因子来操纵与神经突触功能及调控相关基因的表达,从而间接的对神经功能造成影响。当MeCP2基因因突变或者缺失丧失功能时,可导致Rett syndrome;而当MeCP2的基因拷贝数异常增多时,却又会导致患者具有严重自闭症症状的疾病,被命名为MeCP2倍增综合症(MECP2 duplication syndrome)。由此可见,体内MeCP2这个基因的表达量如同一座天平,必须保持着精妙的平衡,无论过多或者过少,都会导致神经突触及神经系统功能的异常,从而导致自闭症。
然而,仅仅知道了遗传背景和分子机制,对于治疗自闭症还是远远不够的。如何为发病过程提供一个观察窗口以及为治疗方案提供一个可供测试的动物模型也是非常重要的,近年来基因编辑技术及模式动物的发展对此提供了非常有利的条件。现有的研究结果已经报道了MeCP2基因敲除的小鼠模型,该模型的确展现出与Rett syndrome非常相似的表型,为Rett syndrome的观察和治疗提供了便利;同时,MeCP2转基因的小鼠模型也同样已得到建立,该模型也展现出明显的社交功能障碍。但自闭症毕竟涉及多种复杂的高级神经活动,如果要为临床研究与治疗提供更为有力的支持,小鼠已显得捉襟见肘,更高级的动物模型和更复杂的神经系统是非常必要的。
为了解决这个问题,中国科学院上海神经科学研究所的科学家们通过使用慢病毒载体侵染带入外源基因的方法(图4A),成功建立了在神经系统中特异性过表达MeCP2基因的转基因食蟹猴模型(图4B)。通过分子生物学和生物化学方法的鉴定,外源的MeCP2基因有效地插入了食蟹猴的基因组之中(图4C),并且能够只在神经系统中进行表达。
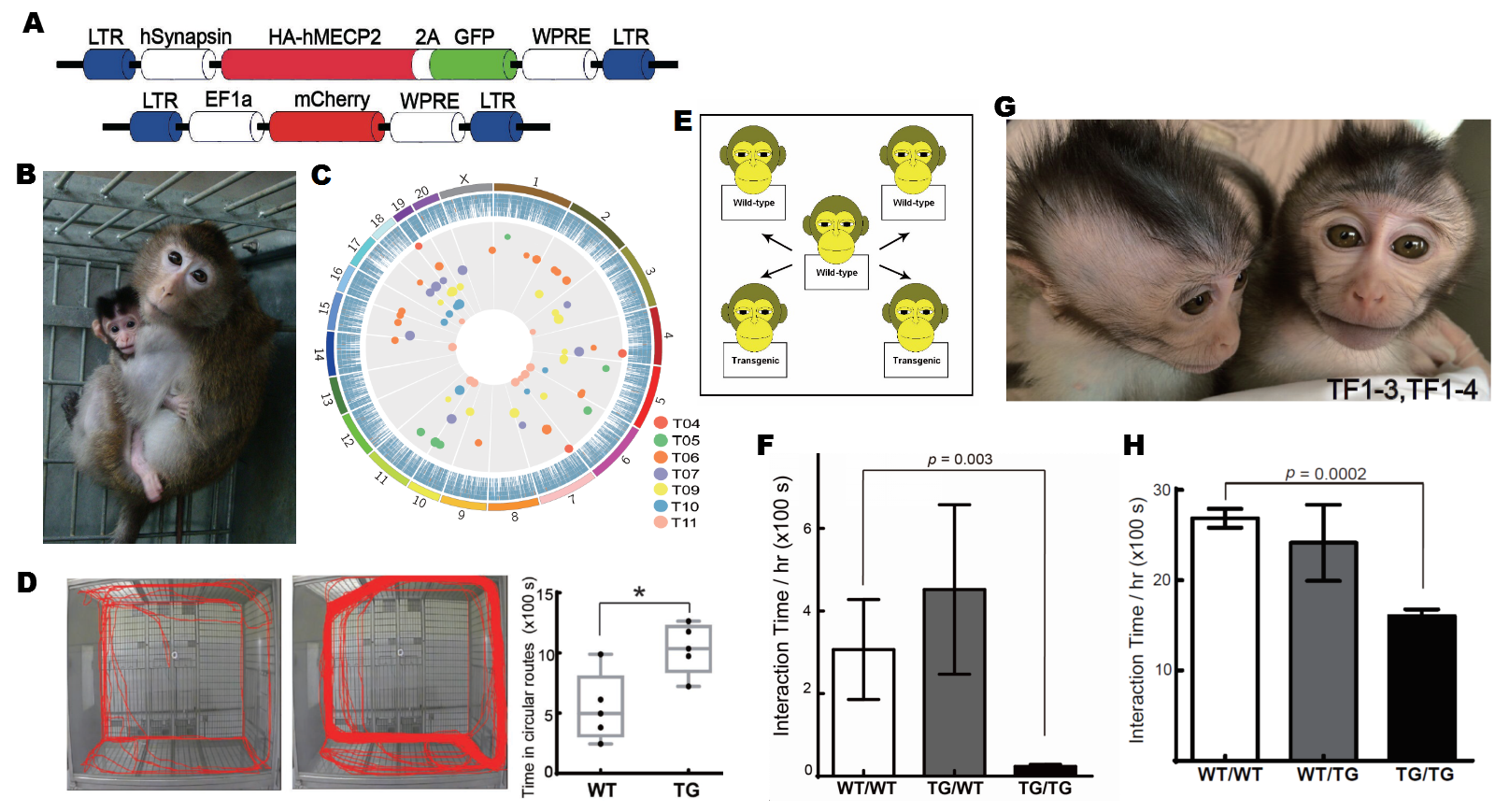
图4 (A)病毒载体中外源转基因片段的设计方案。(B)在母亲照看下的新生F0代转基因食蟹猴。(C)深度测序得到的转基因插入位点的全基因组Mapping。(D)行动路线追踪结果示意及重复路径时间结果统计。(E)社群内社交行为检测的行为学范式。(F)F0代配对社交行为测试的结果。(G)新生的F1代转基因食蟹猴。(H)F1代配对社交行为测试结果。
遗传学层面的操作已经成功,种子既已种下,会在食蟹猴这一非人灵长类动物的土壤中结出怎样的果实呢?转基因食蟹猴相对于野生型的对照食蟹猴又会展现出怎样的不同呢?带着这样的疑问,研究人员们对此模型进行了长期的体征观察和大量的行为学测试。在体征观察与记录中,MeCP2转基因食蟹猴表现出明显的体重发育迟缓以及血浆中脂肪酸代谢的异常,这两种表型也见于一些自闭症患者的临床表现的报道中。而在行为方面,对食蟹猴进行行动路线追踪的结果显示,转基因食蟹猴会花费更多的时间在一种刻板的重复性路径行进上(图4D)。而当人为的给予转基因食蟹猴威胁性刺激时,相对于野生型食蟹猴,该型食蟹猴会表现出更强烈的焦虑情绪与敌意。在最为重要的社交行为及能力方面,研究人员发现,无论是在社群内的社交行为(图4E)还是与另一只配对的社交行为,转基因食蟹猴的社交频率统计结果均显著低于野生型对照(图4F),该结果对于评估MeCP2转基因食蟹猴的类自闭症表型具有重要的意义。在此之后,研究人员又通过威斯康星认知行为测试对转基因食蟹猴的学习能力进行了评估,在该项实验中,转基因食蟹猴并没有表现出明显的学习障碍,但却出现了被认为是自闭症表征的重复刻板行为。
在自闭症中,遗传是一项重要因素,为了深入研究此问题,在研究人员的努力下,通过将转基因食蟹猴的精巢组织移植至裸鼠皮下发育成熟,成功地在短期内得到了原代转基因食蟹猴(F0)的子代猴(F1)(图4G),经过遗传学检测,研究人员们发现,F0代转基因食蟹猴基因组中的外源插入片段的位点,通过种系传递,遗传至F1代子猴的基因组中,并且F1代子猴中位点的分布与F0代比较,呈现出典型的孟德尔分离现象。此结果显示,F1子代猴从亲代F0身上遗传到了神经系统中特异性过表达MeCP2的特性,并且针对于F1的配对社交行为检测结果也显示,转基因食蟹猴相较于野生型对照也展现出社交行为频率的显著下降(图4H),该表型与F0亲代猴以及自闭症患者的临床表型是相似的。
在此项研究中,科学家成功建立了神经系统特异性表达MeCP2的转基因食蟹猴模型,且该转基因可以通过种系传递向子代进行遗传。并且,在第一代与第二代转基因猴中,MeCP2的异常表达都导致了转基因食蟹猴出现明显的类自闭症行为,简而言之,使猴子患上了自闭症。该项工作为观察自闭症的神经科学机理研究提供了一扇重要的窗口,并为之后进一步的研究自闭症的干预方法提供了坚实的基础,最终为自闭症的临床治疗提供了良好的动物模型和试验平台。在接下来的工作中,研究者已经开展了对患有自闭症的MECP2转基因猴进行详细的脑成像研究,以期待用基因编辑的工具对这些携带自闭症基因的转基因猴进行基因治疗,探索最终治愈自闭症之路。
参考文献
Liu, Z., Li, X., Zhang, J., Cai, Y., Cheng, T., Cheng, C., Wang, Y., Zhang, C., Nie, Y., Chen, Z., Bian, W., Zhang, L., Xiao, J., Lu, B., Zhang, Y., Zhang, X., Sang, X., Wu, J., Xu, X., Xiong, Z., Zhang, F., Yu, X., Gong, N., Zhou, W., Sun, Q.* & Qiu, Z.*(2016)Autism-like behaviours and germline transmission in transgenic monkeys overexpressing MeCP2. Nature 530:98-102
 附件下载:
附件下载: 